Fatigue

When we think of skeletal muscles getting tired, we often use the word fatigue, however, the physiological causes of fatigue vary considerably. At the simplest level, fatigue is used to describe a condition in which the muscle is no longer able to contract optimally. To make the discussion easier, we will divide fatigue into two broad categories: Central fatigue and peripheral fatigue. Central fatigue describes the uncomfortable feelings that come from being tired, it is often called "psychological fatigue." It has been suggested that central fatigue arises from factors released by the muscle during exercise that signal the brain to "feel" tired. Psychological fatigue precedes peripheral fatigue and occurs well before the muscle fiber can no longer contract. One of the outcomes of training is to learn how to overcome psychological fatigue. As we train, we learn that those feelings are not so bad and that we can continue to perform even when it feels uncomfortable. For this reason, elite athletes hire trainers that push them and force them to move past the psychological fatigue.
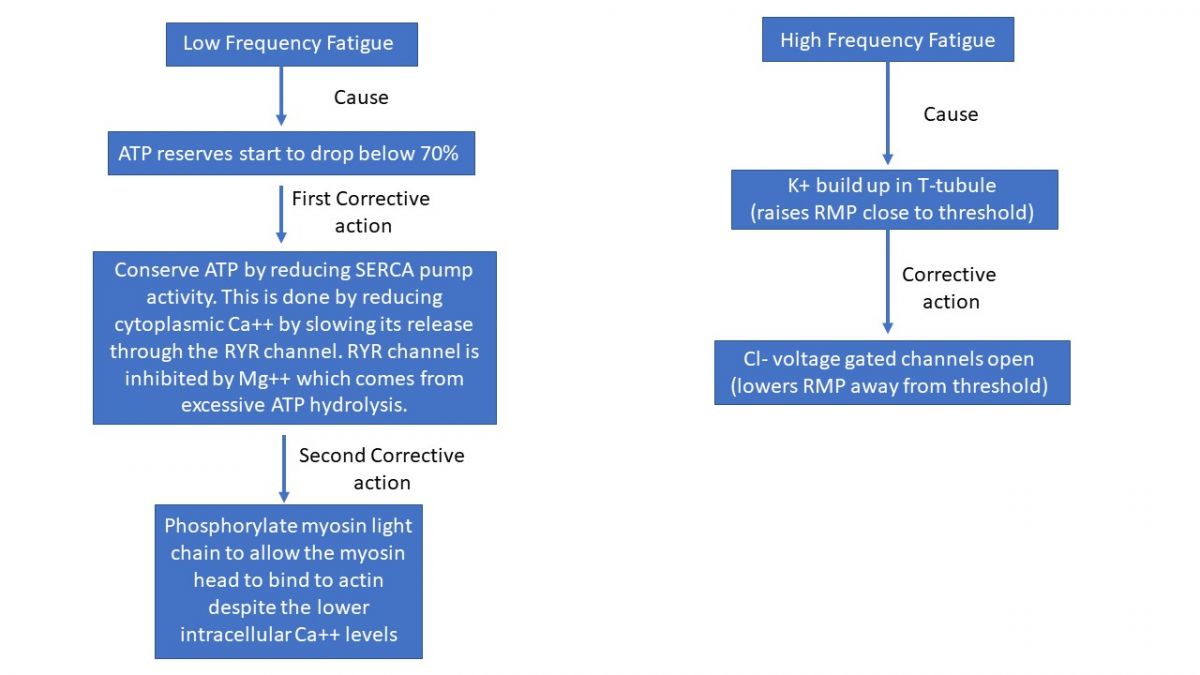
Peripheral fatigue can occur anywhere between the neuromuscular junction and the contractile elements of the muscle. It can be divided into two subcategories, low frequency (marathon running) and high frequency (circuit training) fatigue. High-frequency fatigue results from impaired membrane excitability as a result of imbalances of ions. The list of potential causes include; inadequate functioning of the Na+/K+ pump, subsequent inactivation of Na+ channels and impairment of Ca2+ channels. Muscles can recover quickly, usually within 30 minutes to as little as 30 seconds, following high-frequency fatigue. Low-frequency fatigue is correlated with impaired Ca2+ release, probably due to excitation coupling contraction problems. It is much more difficult to recover from low-frequency fatigue, taking from 24 hours to 72 hours.
In addition, there are many other potential fatigue contributors, these include: accumulation of inorganic phosphates, hydrogen ion accumulation and subsequent pH change, glycogen depletion, and imbalances in K+. Please note that a factor not on the list is lactic acid as it does not contribute to fatigue or muscle soreness. The reality is we still don't know exactly what causes fatigue and much research is currently devoted to this topic.
You should also recognize that a factor not on the list causing fatigue is ATP. This is because experiments performed with muscles show that even in the most severe fatigued conditions of skeletal muscle, 70% of available ATP is still found within the cell! It would appear that ATP is so important for muscle function that other factors will induce fatigue before ATP becomes too low for physiological function. To ensure that ATP is conserved while still maintaining force under constant demand is our next discussion topic.
The current research seems to be supporting the following hypothesis. To conserved ATP without compromising force, the muscle cell must modify the actions of two of the highest consumers of ATP: the SERCA pump and the myosin ATPase enzyme. The SERCA pump can be slowed by reducing the amount of calcium in the sarcoplasm, but reducing the amount of calcium effects myosin’s ability to bind to actin. Thus, the conundrum is the paradox encountered when trying to slow down the SERCA pump with less calcium but without the letting the reduced calcium compromise contraction strength. This is solved by altering the affinity of myosin for actin, but first lets explain how to reduce calcium.
It has been hypothesized that an increase in intracellular Mg2 decreases the responsiveness of the RYR channel. Since Mg2+ is a stabilizing agent for ATP and subsequently released after each hydrolytic reaction, the more intense the contraction, the more Mg2+ will be released. The increase in concentration of Mg2+ will then act to alter RYR, thereby reducing intracellular calcium.
Although not yet well understood, this alteration in calcium triggers the activation of an enzyme complex made up of calmodulin and myosin light chain kinase (MLCK). Once activated, MLCK acts to phosphorylate one of the light chains on the myosin molecule, altering the myosin/actin binding affinity. In short, this modification results in an increase rate of engagement (higher binding affinity for myosin to actin) so that less calcium is required to move troponin. Stated another way, the additional phosphorylation gives the myosin molecule the ability to simply push tropomyosin out of the way. Collectively, these modifications result in the muscle being able to generate force while conserving ATP under conditions of lower intracellular Ca2+.
A second problem with fatigue is the ion imbalance that results from constant depolarization and repolarization. When a muscle membrane is constantly depolarizing and repolarizing the thin diameter of the T-tubule can experience a build-up in extracellular K+. Normal extracellular K+ concentrations range from 3.5 to 5 mM, but under conditions of constant depolarization/repolarization the concentrations can rise to over 11mM in the T-tubules. This high extracellular concentration in K+ results in a membrane potential that repolarizes to a value well above threshold. This high membrane potential inactivates voltage gated channels making it more difficult to propagate action potentials, leaving the muscle cells with even less calcium. To compensate, the T-tubules have additional proteins channels for the Cl- ion. T-tubules contain both leak and voltage-gated Cl- channels as well as ATP sensitive Cl- channels. At concentrations around 11mM K+, the voltage gated Cl- channels can open and help the membrane repolarize. In fact, the importance of Cl- cannot be overstated as studies show that even under resting conditions, the negative 85mV resting potentials of skeletal muscle cells is the result of Cl- channels (80%) rather than K+ channels (20%). The importance of Cl- channels was discovered from observations and studies of the fainting goat. A species that “faints” when overexcited. We now know that the fainting goat lacks Cl- channels in their T-tubules. Even though muscles will still fatigue, without these compensatory mechanisms they would fatigue even faster.

This content is provided to you freely by BYU-I Books.
Access it online or download it at https://books.byui.edu/bio_461_principles_o/fatigue.